مشكلة التمثيل الغذائي للرضع
تم استخدام الملاحظات الموسعة المنشورة في أوائل التسعينيات لاشتقاق معايير معيارية لعملية التمثيل الغذائي القاعدية للرضع. نظرًا لاعتبارات عملية ، فقد تم استخدام معدل التمثيل الغذائي للنوم (SMR) كبديل لعملية التمثيل الغذائي الأساسي للرضع. تمثل SMR ، في ظل معظم ظروف القياس السائدة ، عمليات نشطة ضرورية للحفاظ على الحياة ، ولكن أيضًا ، بدرجات متفاوتة ، تكلفة الطاقة للنمو ، والتأثير الحراري للتغذية ، وبعض حركة الجسم. في مرحلة الطفولة ، يتم حساب التمثيل الغذائي الأساسي في المقام الأول عن طريق الدماغ والكبد والقلب والكلى.
يعتبر الانخفاض في التمثيل الغذائي الأساسي بالنسبة لوزن الجسم ثانويًا لمعدلات النمو التفاضلية لهذه الأعضاء الحيوية بالنسبة للعضلات والدهون. الأيض القاعدي للرضع يتناسب تقريبًا مع وزن الجسم ، وليس مع القوة الكسرية البالغة 0.75 المحددة للبالغين. في سلسلة من الملاحظات التي أجريت على 105 رضع ، كان SMR مرتبطًا عكسياً بوزن الجسم ومعايير بدانة الجسم. اختلف SMR عن طريق وضع التغذية ، ولكن ليس حسب العمر أو الجنس.
تمثل معلمات الوزن / الطول 2 ، ووضع التغذية والطول ، أو الوزن ، والطول ، ووضع التغذية 78 ٪ من التباين في SMR. في تحليل مجموعة فرعية ، لم تزيد كتلة الجسم الخالية من الدهون من إمكانية التنبؤ بـ SMR.
لم تأخذ التطورات في قياسات SMR في الحسبان تمامًا التباين الفردي الذي لوحظ في التمثيل الغذائي الأساسي للرضع.
التاريخ للمرض
أجرت شركة فورستر (1877) أول تحقيق نُشر عن التمثيل الغذائي الغازي للرضع في ميونيخ. تم قياس إنتاج ثاني أكسيد الكربون لطفلين ، أحدهما يبلغ من العمر 14 يومًا والآخر 16 يومًا ، في مقياس كالوريمتر مفتوح الدائرة.
تم استخدام الجهاز لاحقًا بواسطة RUBNER و HEUBNER (1898) في دراساتهما الكلاسيكية عن متطلبات الطاقة اليومية للرضع العاديين والضمورين.
كان RUBNER (1883) رائدًا لدراسات حول أكسدة الركيزة وصاغ قانون مساحة سطح الجسم ، أي النظرية القائلة بأن معدل التمثيل الغذائي لكل وحدة مساحة سطح إما هو نفسه بالنسبة للحيوانات الكبيرة والصغيرة أو على الأقل مستقل عن حجم الجسم. استكشف العلماء الفرنسيون تأثيرات مساحة سطح الجسم (RICHET ، 1885) ودرجات حرارة البيئة والجسم على إنتاج الحرارة (LANGLOIS ، 1887). تمت دراسة التمثيل الغذائي للرضع (VO2 و VCO2) في وقت واحد في إيطاليا بواسطة MENSI (1894) و POPPI (1900) ، وفي تشيكوسلوفاكيا بواسطة SCHERER (1896) و BABAK (1901 ، 1902) باستخدام مقياس السعرات الحرارية الدائرة المغلقة.
تم إجراء سلسلة رائعة من التجارب من قبل SCHLOSSMANN و OPPENHEIMER و MURSCHHAUSER (1908) في دوسلدورف.
أدرك هؤلاء العلماء أهمية الراحة العضلية وتناول الطعام في قياس التمثيل الغذائي الأساسي.
في أمريكا ، بدأ Atwater and Lusk العديد من التحقيقات وقدمت HOWLAND (1911) مساهمة مهمة من خلال إظهار تكافؤ المسعرات المباشرة وغير المباشرة عند الرضع. في تجاربه ، تراوح الفرق بين الطريقتين من -1٪ إلى 3٪.
تم نشر سلسلة من الاستقصاءات حول حديثي الولادة والخدج والرضع الناضجين من قبل بنديكت وتالبوت بين عامي 1914 و 1938. في مستشفى ماساتشوستس العام ، درسوا 37 رضيعًا ، تتراوح أعمارهم بين يوم واحد و 17 شهرًا ، من حالات غذائية مختلفة (بينيديكت وتالبوت ، 1914).
تحدى بنديكت قانون سطح روبنر.
وخلص إلى أن التمثيل الغذائي الأساسي لا يتحدد بوزن الجسم أو مساحة سطح الجسم ، ولكن بالكتلة النشطة للنسيج البروتوبلازمي. في سلسلة لاحقة من الدراسات ، فحص بنديكت التمثيل الغذائي الأساسي لـ 105 من الأطفال حديثي الولادة (بينيديكت وتالبوت ، 1915). بلغ متوسط إنتاج الحرارة الأدنى 42 كيلو كالوري / كجم / يوم للرضع حديثي الولادة. في التحقيق الأكثر شمولاً ، درس بينيديكت وتالبوت (1921) الحد الأدنى من إنتاج الحرارة لـ 73 رضيعًا من 8 أيام إلى 25 شهرًا. قام MURLIN و HOOBLER (1915) بقياس إنتاج الحرارة لعشرة أطفال تتراوح أعمارهم من 2 إلى 12 شهرًا ؛ بلغ متوسطه 60 كيلوكالوري / كجم / يوم وكان الأعلى عند الأطفال المصابين بالضمور ونقص الوزن والأدنى عند الأطفال الذين يعانون من زيادة الوزن. درس ليفين التمثيل الغذائي الأساسي ومتطلبات الطاقة الإجمالية للرضع العاديين ونقصي الوزن مع التركيز على تأثير الطعام والبكاء والنشاط العضلي (ليفين وآخرون ، 1927 ؛ ليفين ، ويلسون وجوتسكول ، 1928 ؛ ليفين وماربلز ، 1931).
أجرى كلاجيت وهاثواي (1941) دراسة طولية لثمانية أطفال تتراوح أعمارهم بين 5 و 10 أشهر.
2. تعريف اساسيات التمثيل الغذائي
التمثيل الغذائي الأساسي هو الطاقة التي يتم إنفاقها في العمليات الخلوية والأنسجة التي تضمن الحفاظ على الحياة. يتم قياس معدل الأيض الأساسي (BMR) في ظل الظروف القياسية التي يكون فيها الفرد في حالة راحة في بيئة محايدة حراريًا بعد 12 إلى 18 ساعة من الصيام (DUBOIS ، 1936 ؛ BENEDICT ، 1938).
يتم قياس معدل الأيض أثناء الراحة (RMR) في ظل ظروف مماثلة ، باستثناء أن الفرد غير صائم ؛ يتم إجراء القياسات بشكل عام بعد ساعات قليلة من تناول وجبة خفيفة (DURNIN and PASSMORE ، 1967). نظرًا لأن الرضع نادرًا ما يكونون هادئين أثناء الاستيقاظ ، فقد لجأ الباحثون إلى قياس معدل التمثيل الغذائي للرضع النائمين. يُعرَّف معدل التمثيل الغذائي للنوم (SMR) بأنه إنفاق الطاقة أثناء النوم ؛ لم يتفق الباحثون بعد على تعريفات لمتغيرات مثل الوقت منذ التغذية وحركة الجسم ومرحلة النوم ومدة القياس. تم إجراء بعض القياسات على الرضع الصائمين والمخدرين (KARLBERG ، 1952) ، ولكن أجريت معظم الدراسات على الرضع الذين يتغذون وينامون (بينيديكت وتالبوت ، 1921 ؛ تالبوت ، 1925 ، 1938). يؤدي النوم والمهدئات إلى خفض معدل الأيض الأساسي وزيادة التغذية. حاول بنديكت تقييم التأثير المربك للتغذية على قياس التمثيل الغذائي الأساسي. قام بقياس إنفاق الطاقة 1 و 2.5 و 5 و 9 و 12 و 18 و 21 ساعة بعد التغذية وخلص إلى أن التأثير كان طفيفًا ، إذا كان هناك تأثير على الإطلاق (بينيديكت وتالبوت ، 1914).
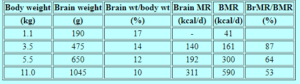
يتم حساب التمثيل الغذائي الأساسي للرضع في المقام الأول عن طريق الدماغ والكبد والقلب والكلى (HOLLIDAY وآخرون ، 1967 ؛ HOLLIDAY ، 1971 ، 1986) (الشكل 1). أظهر تحليل معدل الأيض الأساسي بالنسبة لوزن العضو أن معدل الأيض الأساسي يزداد بمعدل أكبر من وزن العضو خلال فترتي الرحم وما بعد الولادة. يعزى زيادة معدل الأيض الأساسي إلى زيادة النشاط الأنزيمي أثناء الانتقال إلى الحياة خارج الرحم. بعد ذلك ، كان النشاط الأيضي لهذه الأعضاء الحيوية متناسبًا مع كتلتها المتزايدة ؛ كان معدل التمثيل الغذائي للأعضاء / كجم من وزن الجهاز ثابتًا إلى حد ما من الطفولة حتى النضج (الشكل 2). نمط نمو هذه الأعضاء فيما يتعلق بالتغير في وزن الجسم موازٍ لنمط الزيادة في معدل الأيض الأساسي بالنسبة للتغير في وزن الجسم (الشكل 3).
الشكل 1. توزيع معدلات الأيض في الدماغ والكبد والعضلات كنسبة مئوية من إجمالي معدل الأيض الأساسي في أوزان مختلفة من الجسم (HOLLIDAY ، 1986).
1. توزيع معدلات الأيض في الدماغ والكبد والعضلات كنسبة مئوية من إجمالي معدل الأيض الأساسي في أوزان مختلفة من الجسم (HOLLIDAY ، 1986).

الشكل 2. معدل التمثيل الغذائي لوغاريتم الأعضاء (OMR) مقابل المجموع اللوغاريتمي لأوزان الأعضاء (OW) (HOLLIDAY ، 1986).

الشكل 3. العلاقة الموازية بين BMR مقابل وزن الجسم ومجموع أوزان الأعضاء مقابل وزن الجسم (HOLLIDAY ، 1986).

مساهمة الدماغ في التمثيل الغذائي الأساسي مرتفعة بشكل استثنائي في فترة حديثي الولادة (87٪) وطوال السنة الأولى من العمر (53 إلى 64٪) (HOLLIDAY ، 1986) (الجدول 1). هناك عمليتان كيميائيتان أساسيتان ، وهما تخليق البروتين وضخ الأيونات ، يقدر أنهما يمثلان 2/3 أو أكثر من التمثيل الغذائي الأساسي (GRANDE ، 1980). تدعم الطاقة التي يستهلكها الدماغ بشكل أساسي النقل النشط للأيونات للحفاظ على واستعادة إمكانات الغشاء التي يتم تفريغها أثناء عملية الإثارة والتوصيل.
الجدول 1. حجم الدماغ ومتطلبات الطاقة بالنسبة إلى وزن الجسم ومعدل الأيض الأساسي في مراحل مختلفة من النمو *
* مقتبس من HOLLIDAY، 1986.
يعتبر الانخفاض في التمثيل الغذائي القاعدي بالنسبة لوزن الجسم ثانويًا للتفاوت في معدلات نمو الأعضاء ذات معدلات التمثيل الغذائي المرتفعة (مثل الدماغ والكبد والقلب والكلى) مقارنةً بالأعضاء ذات معدلات التمثيل الغذائي المنخفضة (أي العضلات والدهون) HOLLIDAY ، 1971 ، 1986) (الجدول 2). يتم تغيير تكوين الجسم مع استمرار النمو. يزيد وزن المخ والكبد والقلب والكلى بالتناسب مع وزن الجسم خلال السنة الأولى ؛ يظل مجموع أوزان الأعضاء كنسبة مئوية من وزن الجسم ثابتًا نسبيًا عند 15٪ خلال فترة الرضاعة. تزداد كتلة العضلات من 20٪ عند حديثي الولادة إلى 23٪ عند الرضع بعمر 1.5 سنة. على الرغم من أن الدهون أكثر تباينًا ، فإنها تميل إلى الزيادة من 12٪ عند الولادة إلى 20٪ عند 1.5 سنة.
الجدول 2. تكوين الجسم في مراحل النمو المختلفة *
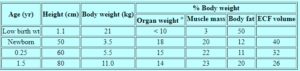
* مقتبس من HOLLIDAY، 1986.
+ مجموع المخ والكبد والقلب والكلى.
في معظم دراسات التمثيل الغذائي القاعدي ، ارتبط قياسه بشكل من أشكال قياس وزن الجسم. اقترح RUBNER (1883) أن استقلاب الطاقة يتناسب مع مساحة سطح الجسم. وفقًا لقانون روبنر ، كان معدل الأيض اليومي (بالكيلو كالوري) يساوي ما يقرب من 1000 ضعف عدد الأمتار المربعة من مساحة السطح. وجد أن هذا غير صالح للأطفال الرضع (بينيديكت وتالبوت ، 1914 ، 1921 ؛ سينكلير ، سكوبس وسيلفرمان ، 1967). ربط المحققون اللاحقون الأيض الأساسي بالقوى الكسرية لوزن الجسم.
درس كليبر (1947 ، 1975) العلاقة بين معدل التمثيل الغذائي (MR) ووزن الجسم (WT) على النحو التالي:
تسجيل الدخول (MR) = تسجيل الدخول (ك) + ن سجل (بالوزن).
أظهر الارتباط الخطي بين السجل MR و log WT أن معدل التمثيل الغذائي كان متناسبًا مع قوة معينة من وزن الجسم. تم العثور على أن الأسس الأنسب لوزن الجسم فيما يتعلق باستقلاب الطاقة هو 0.75 عبر العديد من الأنواع. في ظل الظروف القياسية ، وجد أن معدل التمثيل الغذائي للحرارة المنزلية البالغة من الفئران إلى الماشية يبلغ متوسط 70 كيلو كالوري لكل كيلوغرام WT0.75 في اليوم. على الرغم من عدم تضمين الرضع في تحقيقات كليبر ، فقد تم تطبيق هذه المعادلة على الرضع.
طبق Karlberg نهج Kleiber لتحليل السجل على بياناته وغيرها من البيانات المنشورة ووجد أن معدل الأيض الأساسي للرضع كان متناسبًا إلى حد ما مع وزن الجسم. كانت الأسس الناتجة عن وزن الجسم 0.918 (KARLBERG ، 1952) ، 1.090 (BENEDICT و TALBOT ، 1921) ، 1.113 (JANET and BOCHET ، 1933) ، و 0.912 (BAER ، 1929). اختلفت القيم بشكل كبير عن 0.75 لكليبر. خلال مراحل النمو السريع ، قد لا يكون 0.75 المشتق من الحيوانات البالغة مناسبًا للرضع. يتوافق إنفاق الطاقة على الصيانة مع الطاقة التي ، وفقًا لكليبر ، مرتبطة بقوة الوزن البالغة 0.75. قد يكون الأس أكبر من 0.75 عند الرضع ، بسبب متطلبات الطاقة الإضافية للنمو ، أو بسبب النشاط الأيضي العالي للأنسجة الدهنية عند الرضع مقارنة بالبالغين. يميل الأس إلى الانخفاض مع انخفاض النمو عند الأطفال الأكبر سنًا.
3. العوامل التي قد تؤثر على التمثيل الغذائي الأساسي
عمر. لا يوجد إجماع حول تأثير العمر على التمثيل الغذائي الأساسي ، الذي يتم تطبيعه حسب وزن الجسم ، خلال السنوات الأولى من العمر. أشارت ملاحظات BENEDICT و TALBOT (1921) إلى أن التمثيل الغذائي الأساسي (kcal / kg / d) زاد خلال أول 12 إلى 18 شهرًا ثم انخفض بعد ذلك. في دراسة طولية أجراها CLAGETT و HATHAWAY (1941) ، كان الإنفاق الأساسي (لكل كيلوغرام) ثابتًا إلى حد ما للأفراد طوال سلسلة الملاحظات. أشارت بيانات KARLBERG (1952) إلى أن معدل الأيض الأساسي لكل كيلوغرام من وزن الجسم انخفض تدريجياً عند الرضع بين أسبوع واحد وسنة واحدة (الشكل 4). أكد جميع الباحثين على الاختلاف الواضح بين المنحنيات الفردية لعملية التمثيل الغذائي القاعدي مقابل العمر.
الشكل 4. العلاقة بين معدل التمثيل الغذائي للنوم (SMR) (kcal / kg / d) والعمر (wks) (KARLBERG ، 1952).

الجنس.
لم يثبت أن الجنس مرتبط باختلافات التأثير في التمثيل الغذائي الأساسي للرضع (بينيديكت وتالبوت ، 1921 ؛ تالبوت ، 1925).
الأصل العرقي.
لم يتم إثبات الاختلافات العرقية في معدل الأيض الأساسي ، غير المرتبط بالحالة التغذوية أو ربما بالظروف المناخية (استشارة خبراء منظمة الأغذية والزراعة / منظمة الصحة العالمية / جامعة الأمم المتحدة ، 1985). القليل من المعلومات المتاحة عن الرضع.
نقص التغذية.
يُعتقد أن معدل الأيض الأساسي لدى البالغين الذين يعانون من نقص التغذية مصاب بالاكتئاب (KEYS وآخرون ، 1950) ؛ التقارير الواردة في الأدبيات أقل وضوحًا فيما يتعلق بالرضع والأطفال. تم الإبلاغ عن انخفاض معدلات التمثيل الغذائي لدى الأطفال المصابين بسوء التغذية الحاد (PARRA et al. ، 1973 ؛ BROOKE ، COCKS and MARCH ، 1974). يبدو هذا الانخفاض متسقًا مع الملاحظات السريرية لانخفاض درجة حرارة الجسم ، وعلى مستوى الأنسجة ، ضعف آليات إنتاج الطاقة في العضلات والفسفرة التأكسدية (WATERLOW and ALLEYNE ، 1971). التقارير المتناقضة للزيادة (بينيديكت وتالبوت ، 1914 ؛ مورلين وهوبلر ، 1915 ؛ ليفين ، ويلسون وجوت تشال ، 1928 ؛ فليمينج وهوتشيسون ، 1924) أو طبيعية (مونتجومري ، 1962 ؛ كريجر وويتن ، 1969) قد تكون معدلات التمثيل الغذائي هي النتيجة القياسات التي أجريت في مراحل مختلفة من التعافي من سوء التغذية.
طعام.
تم الخلط بين معظم قياسات التمثيل الغذائي الأساسي للرضع بسبب التأثير الحراري للتغذية (TEF) ، والذي حاول العديد من الباحثين قياسه (HOWLAND ، 1911 ؛ بينديكت وتالبوت ، 1921 ؛ مورلين ، 1925 ؛ ليفين ، 1927). قدّر بنديكت أن TEF أعلى بنسبة 8 إلى 15٪ من النفقات الأساسية ، لكنه أشار أيضًا إلى الطابع الفردي الواضح للاستجابة. في ملاحظاته ، التي استمرت حتى 11 ساعة بعد الرضاعة ، لم يتم العثور على الحد الأدنى المطلق من TEF عادة قبل 8 إلى 9 ساعات بعد الأكل. كان تأثير TEF على التمثيل الغذائي الأساسي يعتمد على حجم التغذية ؛ مع تغذية صغيرة (حوالي 50 كيلو كالوري) ، اقتربت القياسات من المستوى الأساسي في نهاية 4 ساعات.
البرد.
يتم تحفيز التوليد الحراري عند الرضع المعرضين لدرجات حرارة محيطة أقل من درجة الحرارة الحرجة. أوضح BRUCK (1961) أنه يمكن زيادة معدل الأيض للرضع حديثي الولادة بنسبة 100٪ تقريبًا استجابة للبرد.
النوم.
ينخفض إنفاق الطاقة مع النوم ويختلف بين مرحلتي حركة العين غير السريعة (NREM) وحركة العين السريعة (REM).
أظهرت الدراسات التي أجريت على الرضع أن VO2 يزداد بنسبة 16٪ (SCOPES و AHMED ، 1966) و 9.4٪ (KAIRAM وآخرون ، 1979) في الانتقال من مراحل حركة العين غير السريعة إلى حركة العين السريعة. يتم تحرير هرمون النمو البشري (hGH) أثناء النوم (SASSIN et al. ، 1969).
ترتبط ارتفاعات هرمون النمو مع تطور نوم حركة العين غير السريعة. بسبب عمل تحفيز النمو لهرمون النمو ، من المحتمل أن تتضمن قياسات SMR تكلفة الطاقة للنمو.
مدة الحمل والنمو داخل الرحم.
معدلات الأيض القاعدية للخدج أقل من معدلات الأيض عند الرضع الناضجين وتزداد بمعدلات أبطأ خلال الشهر الأول من الحياة خارج الرحم. عند الحياد الحراري ، لم تتجاوز معدلات التمثيل الغذائي للخدج (1000-2000 جم ، من 2 إلى 31 يومًا من العمر) 40 كيلو كالوري / كجم / يوم (MESTYAN و JARAI و FEKETE ، 1968). نظرًا لحجم دماغهم الكبير نسبيًا ومعدلات نموهم المرتفعة ، يكون استهلاك الأكسجين أعلى في عمر الحمل الصغير (SGA) مقارنة بالخدج المناسبين لسن الحمل (AGA) (SINCLAIR and SILVERMAN ، 1964).
المصدر:ترجم لموقع اجنبي
هذا الرابط للموقع
https://archive.unu.edu/unupress/food2/UID09E/UID09E0E.HTM#:~:text=The%20basal%20metabolism%20of%20infants,not%20by%20age%20or%20sex.



